El Metabolismo Autótrofo
Los autótrofos son, en las cadenas tróficas, los productores, y son el primer eslabón de todas ellas, pues son los organismos que convierten el CO2 y demás materia inorgánica en materia orgánica.
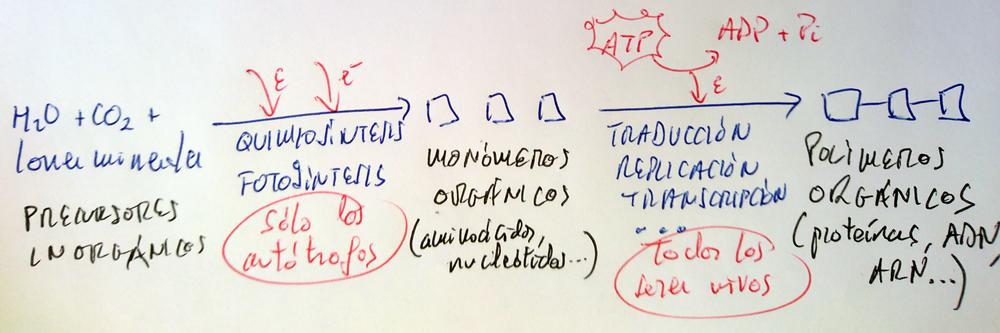
Este proceso de transformación de la materia inorgánica en orgánica consume energía y electrones (poder reductor).
Tipos de Metabolismo Autótrofo
Según cuál sea la fuente de energía, los autótrofos pueden ser:
- Los fotoautótrofos realizan la fotosíntesis: producen materia orgánica a partir de CO2 y de la energía de la luz.
- Los quimioautótrofos litótrofos realizan la quimiosíntesis: producen materia orgánica a partir de CO2 y de la energía de enlace químico de sustancias inorgánicas.
Además, la fuente de electrones también puede variar. De todo ello resultan 3 tipos básicos de metabolismo autótrofo:
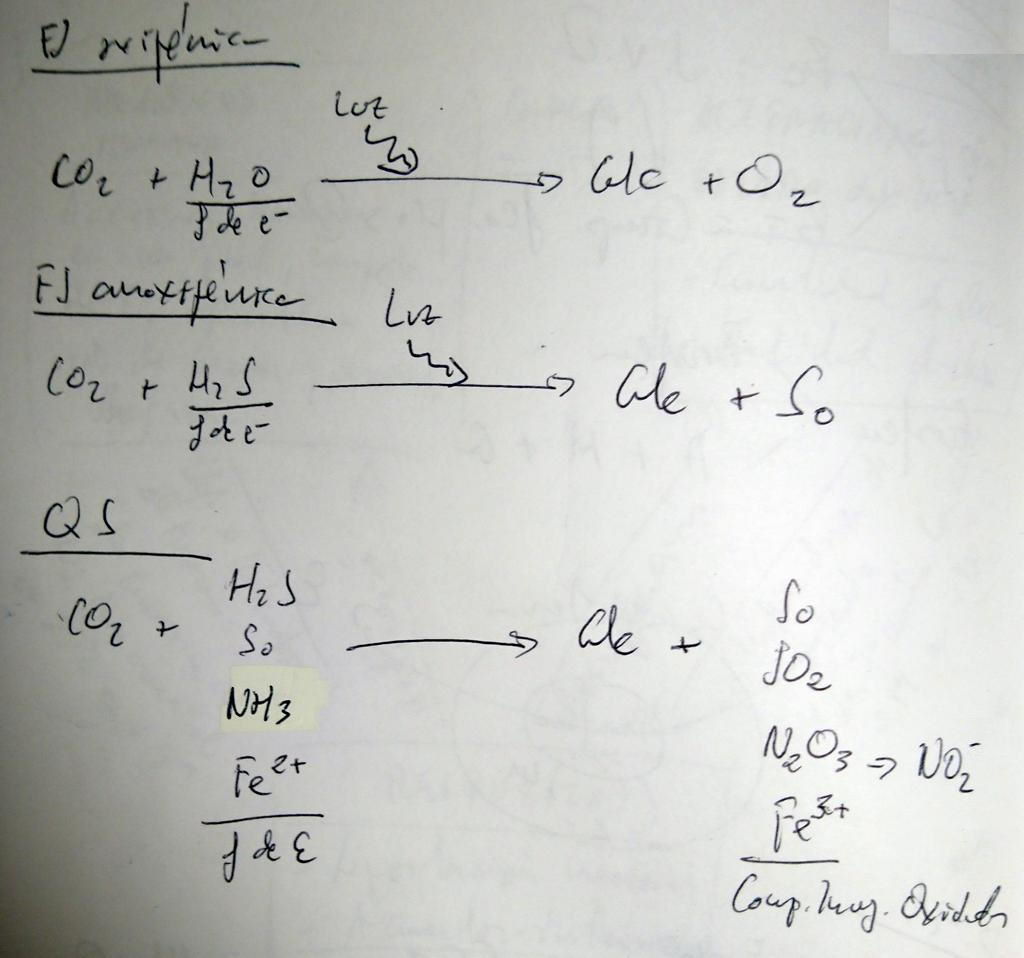
- Fotosíntesis oxigénica (la fuente de e- es el H2O):
- Cianobacterias;
- Algas;
- Plantas.
- Fotosíntesis anoxigénica (la fuente de e- es el H2S):
- Bacterias verdes y púrpuras del S.
- Qumiosíntesis (la fuente de e- son las mismas moléculas inorgánicas que aportan la energía):
- Arqueas;
- Bacterias nitrificantes;
- Otras bacterias oxidantes (del S, del Fe, del H2...).
La Fotosíntesis Oxigénica
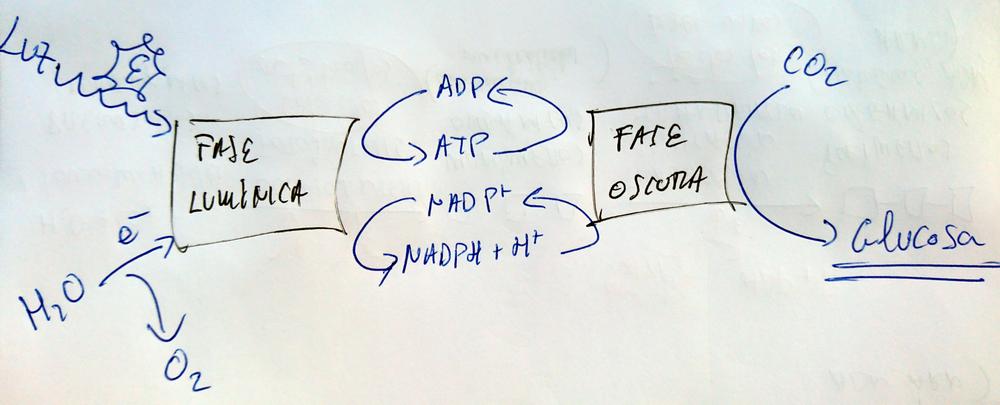
Etapas
- Fase lumínica.
- Captación de la energía lumínica.
- Cadena transportadora de electrones
- abierta (acíclica);
- cerrada (cíclica).
- Fotofosforilación.
- Fase oscura.
- Ciclo de Calvin.
Fase Lumínica
Sucede en los tilacoides de los cloroplastos.
Captación de la energía lumínica
- Por pigmentos fotosintéticos.
- En algas verdes y plantas:
- Clorofilas a y b: son cromoproteínas con un fitol (lípido isoprenoide del grupo de los diterpenos) y un anillo porfirínico con Mg2+ en el centro.
- Carotenoides: son lípidos isoprenoides del grupo de los tetraterpenos:
- ß-caroteno (naranja);
- xantofilas (amarillas);
- licopeno (rojo).
- Agrupados en los fotosistemas PSI y PSII, con:
- Complejo antena: pigmentos captadores de la luz;
- Centro de reacción: una única molécula de clorofila llamada P700 (en el PSI) o P680 (en el PSII), según la longitud de onda a la que capten el máximo de luz.
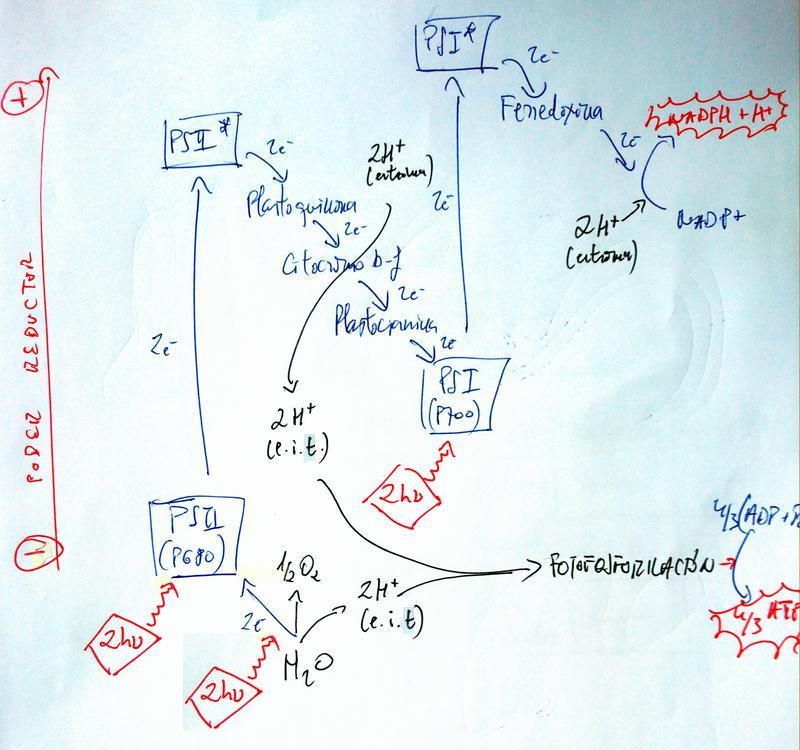
Flujo electrónico abierto (acíclico) y fotofosforilación
- Un par de fotones inciden en el PSI, cuya clorofila P700 pierde 2 e- al ser excitados a un nivel energético superior y saltar de la molécula.
- Estos 2 e- caen por una cadena transportadora hasta un aceptor final, el NADP+, que se reduce a NADPH + H+.
- Un par de fotones inciden en el PSII, cuya clorofila P680 pierde 2 e- al ser excitados a un nivel energético superior y saltar de la molécula.
- Estos 2 e- caen por una cadena transportadora hasta llegar a la clorofila P700 del PSI, que recupera así los 2 e- que había perdido. En este flujo se desprende energía que permite el bombeo de 2 H+ desde el estroma del cloroplasto al interior de los tilacoides, donde se acumulan en contra de gradiente.
- Un par de fotones inciden en una molécula de H2O, que se disocia en:
- 1/2 O2;
- 2 e-, que reponen los perdidos por el PSII; y
- 2 H+, que se acumulan en el interior de los tilacoides en contra de gradiente.
- Los 4 H+ acumulados en el espacio intratilacoidal retornan al estroma del cloroplasto a favor de gradiente a través de una ATP-sintasa. Por cada 3 H+ que pasan se produce 1 ATP a partir de ADP y Pi libres en el estroma. En total se producen, por tanto, 4/3 ATP. Este proceso se llama fotofosforilación.
- En conjunto:
- Hay un flujo de 2 e- desde el H2O (donador de e-) hasta el NADPH (aceptor final de e-) causado por la incidencia de los fotones.
- La energía de los fotones va a parar al ATP.
- Balance:
Entradas Salidas Materia 1 H2O 1/2 O2 Poder reductor 1 NADP+ 1 NADPH + H+ Energía 6 fotones 4/3 ATP
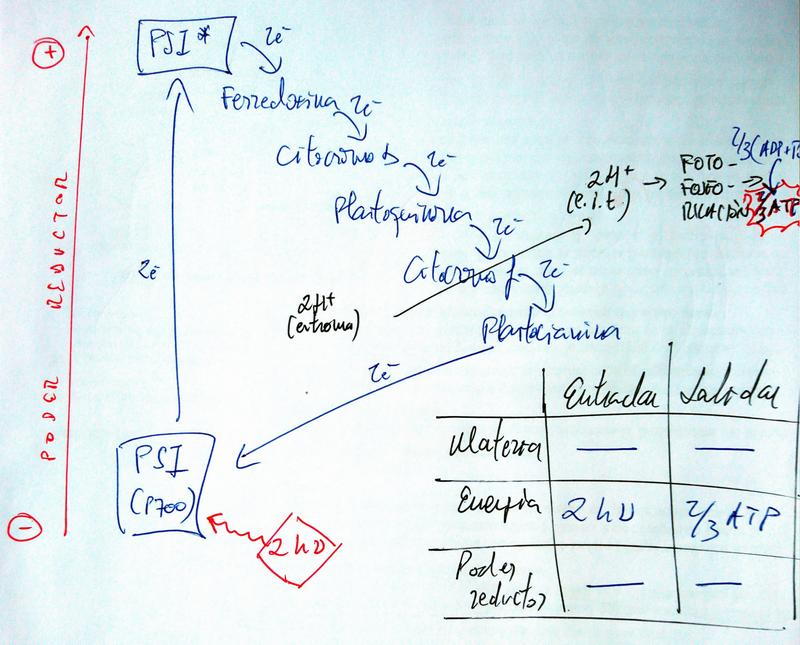
Flujo electrónico cerrado (cíclico) y fotofosforilación
- Sólo interviene un fotosistema: el PSI.
- El flujo de e- es desde el PSI al PSI.
- Por tanto, el aceptor final de e- no es el NADP+ y no se genera poder reductor.
- Tampoco hay entradas ni salidas de materia.
- Sirve, únicamente, para producir ATP.
- Como solo se bombean 2 H+ al espacio intratilacoidal, solo se producen 2/3 de ATP por fotofosforilación en cada vuelta.
- La realizan las células verdes cuando necesitan producir ATP, pero no poder reductor (por ejemplo para compensar los ATP que hacen falta para la fase oscura y que no se producen en la fotofosforilación abierta; o para complementar la respiración celular).
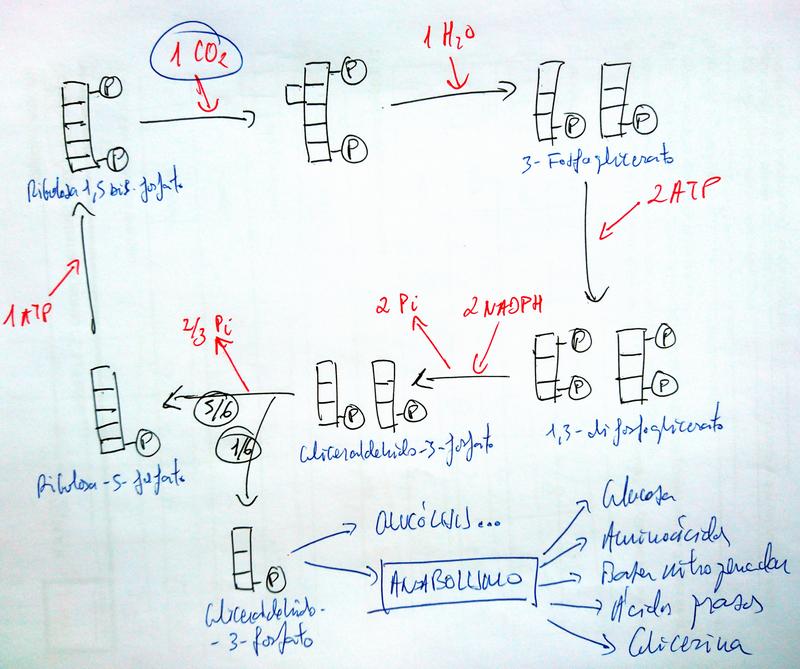
Fase Oscura o Ciclo de Calvin
- Tiene lugar en el estroma de los cloroplastos.
- En ella se consumen el ATP y el NADPH producidos en la fase lumínica, para poder producir materia orgánica a partir de la inorgánica.
- El proceso comienza cuando la enzima RuBisCO captura una molécula de CO2 y la fija a la ribulosa-1,5-bisfosfato. Y termina con la regeneración de esta misma molécula.
- El rendimiento es una molécula de gliceraldehído-3-fosfato (GA3P) cada 3 vueltas (el GA3P tiene 3 átomos de carbono, y en cada vuelta solo se fija 1).
- Luego, ulteriores procesos anabólicos, que pueden necesitar o no de los iones minerales que ascienden por la savia bruta, pueden utilizar esta molécula para producir...
- glucosa, que se podrá utilizar en la síntesis de todos los demás glúcidos;
- aminoácidos, que se podrán utilizar en la síntesis de proteínas;
- bases nitrogenadas orgánicas, que se podrán utilizar en la síntesis de nucleótidos, y por tanto de ácidos nucleicos;
- glicerina y ácidos grasos, que se podrán utilizar en la síntesis de acilglicéridos y otros lípidos;
- factores de crecimiento, hormonas vegetales, ...
- Pero también se podrá consumir en la glucólisis, como primer paso de la respiración celular para producir ATP.
- Luego, ulteriores procesos anabólicos, que pueden necesitar o no de los iones minerales que ascienden por la savia bruta, pueden utilizar esta molécula para producir...
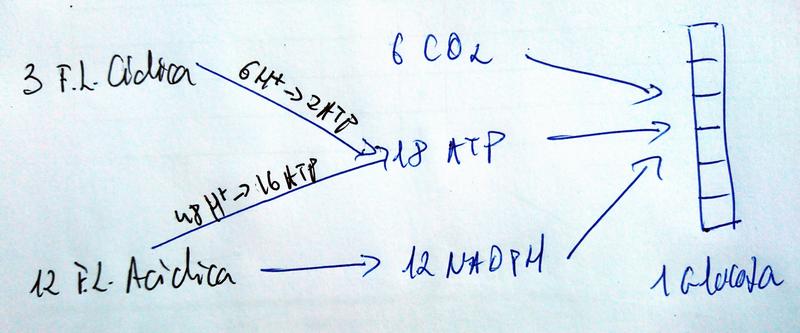
Balance Global de la Fotosíntesis en referencia a una molécula de Glucosa
- Como en cada vuelta del ciclo de Calvin solo se fija 1 C y la glucosa tiene 6, pues hacen falta 6 vueltas para poder sintetizar una molécula de glucosa.
- Como en cada vuelta se gastan 3 ATP y 2 NADPH, pues se necesitan 18 ATP y 12 NADPH para poder sintetizar una molécula de glucosa.
Entradas:
1 vuelta 3 vueltas 6 vueltas CO2 1 3 6 H2O 1 3 6 ATP 3 9 18 NADPH 2 6 12 Salidas:
1 vuelta 3 vueltas 6 vueltas GA-3-Fosfato 1/3 1 2 NADP+ 2 6 12 ADP 3 9 18 Pi 8/3 8 16 Para obtener esos 12 NADPH basta con 12 ejecuciones de la fase lumínica abierta.
- Pero como cada una solo produce 4/3 de ATP, pues esas 12 ejecuciones solo rinden 16 ATP. Y hacen falta 18.
- Los 2 que faltan se obtienen con 3 vueltas a la fase lumínica cerrada, ya que cada vuelta rinde 2/3 de ATP.
Factores que Condicionan la Tasa de Actividad Fotosintética
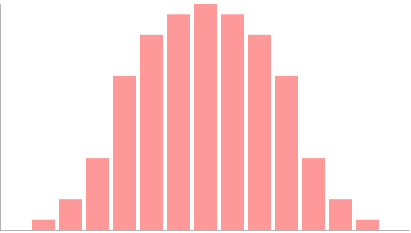
Temperatura
A mayor temperatura, mayor agitación de las moléculas implicadas en la fotosíntesis, mayor probabilidad de encuentro entre ellas, y por tanto, mayor tasa de actividad fotosintética. Hasta un óptimo, a partir del cual las proteínas comienzan a desnaturalizarse y/o las plantas cierran sus estomas para evitar deshidratarse. A partir de ese óptimo la tasa de actividad fotosintética disminuye.
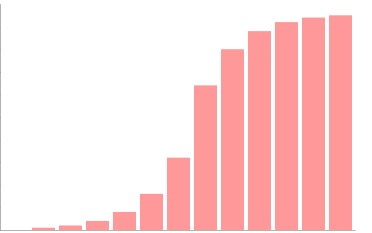
Intensidad luminosa
A mayor intensidad luminosa, mayor tasa de fotólisis del agua y de excitación de los electrones del centro de reacción de los fotosistemas, y por tanto, mayor tasa de actividad fotosintética. Hasta un óptimo, a partir del cual no hay más fotosistemas cuyos centros de reacción se puedan excitar. Ese óptimo depende de la especie, lo que permite que haya tanto plantas de penumbra, como otras que precisan de una elevada intensidad luminosa.
Humedad relativa
A mayor humedad relativa, mayor tasa de apertura estomática (ya que disminuye el riesgo de que la planta se deshidrate), mayor ritmo de entrada de CO2 y de salida de O2, y por tanto, mayor tasa de actividad fotosintética. Hasta un óptimo, a partir del cual todos los estomas están abiertos al máximo.
Concentración de CO2
A mayor concentración de CO2, mayor tasa de fijación de CO2 por la RuBisCO, y por tanto, mayor tasa de actividad fotosintética. Hasta un óptimo, a partir del cual no hay más unidades de RuBisCO libres.
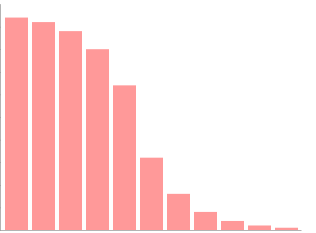
Concentración de O2
A mayor concentración de O2, mayor tasa de fotorrespiración y menor tasa de fijación de CO2 por la RuBisCO, y por tanto, menor tasa de actividad fotosintética.
Longitud de onda de la radiación incidente
Existen longitudes de onda menos eficaces, como la de la luz de color verde, que es menos absorbida por el conjunto de pigmentos fotosintéticos de las plantas.
Es necesario, además, que la radiación incidente incluya radiaciones de 680 nm y 700 nm de longitud de onda para que puedan excitarse los fotosistemas II y I, respectivamente.
La Quimiosíntesis
- La realizan solo las arqueas y algunas bacterias.
- Son organismos quimioautótrofos litótrofos. Obtienen el C del CO2 y la energía de moléculas inorgánicas reducidas.
- De entre las formas de ser autótrofo, la quimiosíntesis requiere una maquinaria enzimática mucho más simple que la fotosíntesis, por lo que se piensa que aquel es un tipo de metabolismo más antiguo. También es importante porque los organismos quimiosintéticos son la base de cadenas tróficas actuales en ecosistemas a los que no llega la luz (fondos oceánicos, lagos subglaciales de la Antártida, y quizás bajo los hielos de Europa y Titán, satélites de Júpiter y Saturno, respectivamente).
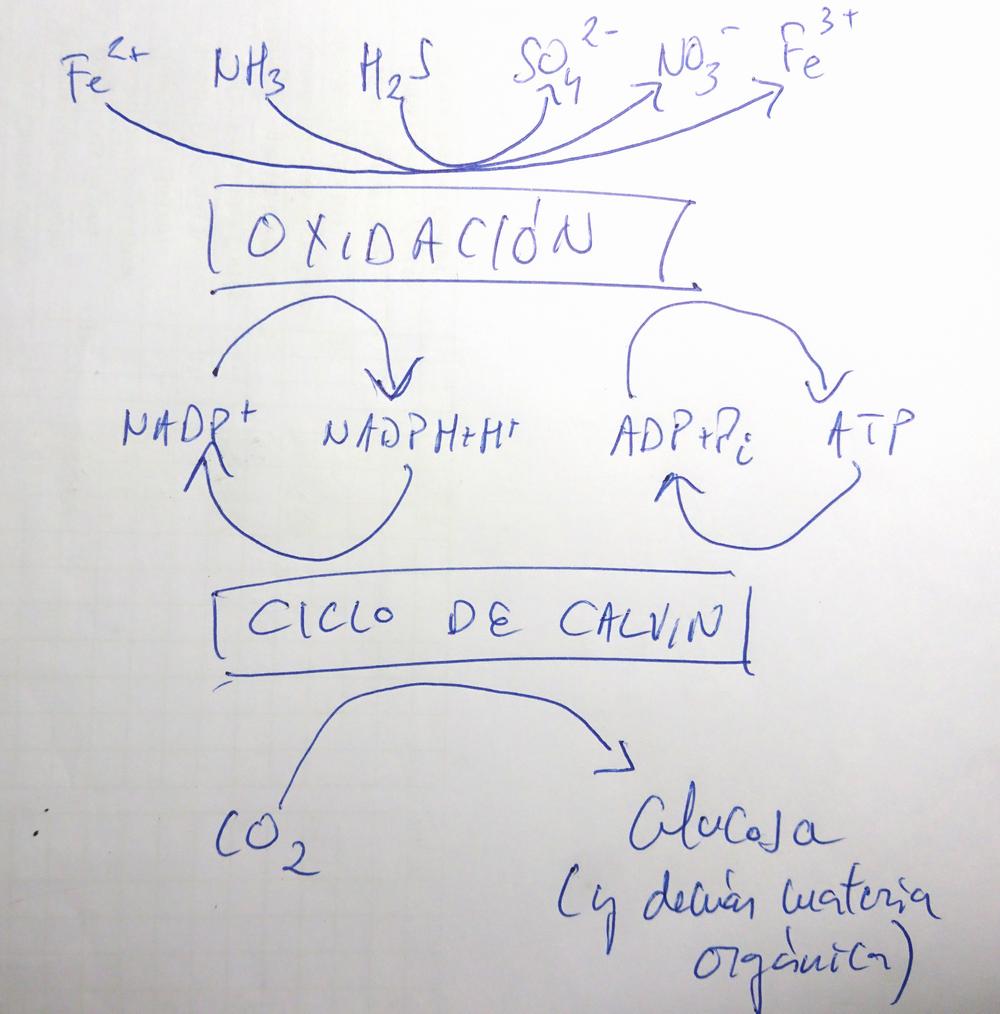
- La quimiosíntesis tiene 2 fases:
- Una etapa de Oxidación. En ella se obtiene ATP y NADPH gracias a la oxidación de moléculas inorgánicas reducidas:
- Bacterias nitrificantes: NH4+ (amonio) → NO2- (nitrito) → NO3- (nitrato).
- Bacterias sulfooxidantes: SH2 → S (azufre elemental) → SO32- (sulfito) → SO42- (sulfato).
- Bacterias ferrooxidantes: Fe2+ (hierro ferroso) → Fe3+ (hierro férrico).
- Bacterias oxidantes del hidrógeno: H2 → H2O.
- El Ciclo de Calvin. En él, como en la fotosíntesis, gracias a la energía del ATP y a los electrones del NADPH, el CO2 es convertido en gliceraldehído-3-fosfato, y éste luego en glucosa y demás moléculas orgánicas.
- Una etapa de Oxidación. En ella se obtiene ATP y NADPH gracias a la oxidación de moléculas inorgánicas reducidas: